
For a primer on this blog series and an FAQ, see here.
| Other Episodes |
||||
| Coasts | Deserts | Freshwater | Ice Worlds | Forests |
| Islands | Badlands | Swamps | Oceans | North America |

This episode features advisement by the following paleontology consultants:
- Steve Brusatte
- Alexander Farnsworth
- Scott Hartman
- John Hutchinson
- Robert Spicer
- Paul Valdes
- Mark Witton
- Darren Naish
Scene 1: Somewhere in Mongolia

Baseless speculation
Canyon-living pterosaurs
This scene shows an unnamed species of pterosaur flying up onto a canyon wall where they are shown taking care of their young. Attenborough tells us that these pterosaurs do this every evening.
As you have probably guessed by this far into this series, we have no evidence for this.
Return of “Velociraptor”
The star of this scene is a group of “Velociraptor”. The timeline issues were discussed in the last installment. Now that Velociraptor is playing more than a bit part in the scene, we can find several more issues.
The first one is the animation of the tails in these animals. The tails act like motionless dowels rather than firm attachment points for the caudofemoralis muscle complex. While it is true that dromaeosaurs reduced their tail musculature compared to most other theropods, the caudofemoralis complex was still a sizeable muscle group that contributed a substantial chunk to the mass of their tails (~24%, Persons and Currie 2012). While its function as the primary driver of femoral retraction may be reduced it would have still be a substantial contributor, which would have affected tail motions more than what we are shown in the show.
Much of this scene focuses on highlighting the feathers in Velociraptor and all their many functions aside from flight. It’s shame then that the primary feathers are shown fully extended the entire time. Naish highlights several aspects of feather functions in Velociraptor that could explain feather evolution. However, all of these examples remain hypothetical with little empirical basis to them. Hence the placement in this section. It’s very clear from the Megathread and the Uncovered segment that making a feathery dromaeosaur was a top priority for this scene.
We are shown a small pack of Velociraptor scaring the unnamed pterosaurs into a frenzy and then taking advantage of the chaos to grab a meal. It’s a scene that appears directly inspired from prior BBC nature documentaries. However, as discussed in the last installment, this is also a scene that relies heavily on the assumption that Velociraptor or any dromaeosaur was a pack hunter. Naish states that we have evidence to support group (not pack) hunting in dromaeosaurs, but we really don’t. As I mentioned last time, the closest evidence that we have comes from Ostrom’s Deinonychus and Tenontosaurus discovery, which has largely been abandoned now as evidence for group hunting.
Mostly speculation
Long-flight pterosaurs
Attenborough tells us that these unnamed pterosaurs “could fly for hundreds of miles in a day”. Whereas, we can look at the anatomy of various pterosaurs and get an idea on their potential flight efficiency, making quantified statements such as this goes too far.
Missed opportunity
The “killing” claw
Despite using Velociraptor for this scene and even showing closeups of the sickle claw, we never see the claw in use throughout the series. This is a shame given how well known the claw is. It would have been a good time to offer an alternate take from the classic slashing action and instead show the puncture and grapple hypothesis (Manning et al. 2009; Fowler et al. 2011). It would all still be speculative (even with a talking head), but all that is par for the course with this show anyway, so why not ?
Scene 2: American Southwest (probably)

Artistic anachronism
Tyrannosaurus roars at the…camera man?
This was a weird cinematic choice. So, now the camera crew is physically present here? I’m not a fan of animals making noise without a reason and the only reason why this would happen in this scene is if the camera crew were physically present in this fictional setting.
Baseless speculation
Tyrannosaurus sniffs the air like a dog or a person
This isn’t how sauropsids sniff. We have good experimental and biomechanical data for how sauropsids sniff the air. They don’t do the ballistic pumping of air in and out that we see in mammals (largely driven by their diaphragms). Instead, sauropsids move air in and out of the nasal passages via buccal / gular pumping (McCutcheon 1943; Naifeh et al. 1970; Dial and Schwenk 1996). Whenever you see a turtle, lizard, or crocodile pulsate its throat, it’s sniffing the air. Birds likely do this too, but we have fewer data on bird olfaction in general, so it’s harder to say for certain.
I might be a bit nit picky and harsh here as this happens to be an area I’m very familiar with, but still. If the point of this series to to portray dinosaurs as accurately as possible, then this level of detail is warranted.
Also, drop the moving nostrils thing. It’s unlikely that any dinosaur (save, possibly spinosaurs) could do this.
Tyrannosaurus washes its wounds in the river
This is a level of health maintenance that most of western civilization didn’t do prior to ~150 years ago. Washing wounds is a rare event. It’s very unlikely that a T. rex would have the forethought to consider this. More likely is the wounds clotting up quickly and the animal’s natural immunity keeping it safe.
Useless tail motions
Again, dinosaur tails were effectively “fifth limbs” for the animal’s (third limb for bipeds). The tails should be showing tension as the legs move back and forth instead of just acting like a dead extension jutting from the back end of the animal.
T. rex interactions and…other bits
A female T. rex enters the male’s territory and they roar at each other, then the male shows a submissive posture with a bright-red gular region. This is followed by by some light face fondling and then a G-rated sex scene. We have no data to support any of these interactions. Yes, T. rex mated, but we don’t know any of the specifics there. We especially don’t know about any potential courtship behaviour.
“A clutch of 15”
After the pair of T. rex “do the deed” we are told that the female will lay up to fifteen eggs. This is based on exactly zero data. As I mentioned in the first installment, we don’t have any tyrannosaur nests nor any hatchlings. This entire section is completely fictitious.
Mostly speculation
The killing of an adult Triceratops
We are shown the large male, T. rex after it had apparently battled and killed a large, adult Triceratops horridus. We do have some evidence that Tyrannosaurus rex hunted and ate Triceratops horridus (e.g., Erickson et al. 1996), it’s far less likely that this was done on large, adult animals. It’s possible that this individual was already sick or elderly. We are not told much here so it’s hard to say one way or the other. This scene comes off as possible but not very probable.
The hypersensitive snout of T. rex
Attenborough informs the audience that the facial area of a tyrannosaur is very sensitive to touch. This interpretation came from studies on the neurovascular foramina in the snouts of T. rex and related tyrannosaurs (Carr et al. 2017). I discussed this in depth here, so I’ll just summarize things by saying that most of these foramina likely belonged to blood vessels as opposed to a hyper-arbourized trigeminal nerve system. In short: no, T. rex probably did not have a uniquely sensitive snout.
Scene 3: Mongolian Swamp

Baseless speculation
Hungry hungry Deinocheirus
The star of this scene is Deinocheirus mirificus. We are shown this shaggy animal wading through a marshy swamp as it devours vegetation. Attenborough tells us that the duck-billed snout of D. mirificus is very effective at gathering water plants. Further, our star dino has to eat so much right now because of the lack of food in the dry season. There are a couple of things wrong here. First off, we don’t know about the food requirements or dietary changes that occurred for most dinosaurs, much less this weirdo among theropods. Although, current data does support a herbivorous diet (Lee et al. 2014), we have no idea how much that diet changed during the year. We are told that the duck-bill is really efficient at collecting water plants, but this seems mostly based on how ducks forage. There have been no tests on cropping efficiency in D. mirificus, nor any data on how the chewing cycle worked in this species.
Harassed by ectoparasites
We see swamp ectoparasites (mosquitoes, gnats, etc.) bothering our star dinosaur. To tackle this frustration, D. mirificus goes to a nearby tree and starts scratching itself on it. This is very reminiscent of what bears do in nature documentaries. This was doubtless inspired by bear behaviour, which is a far cry from Sauropsida, so it’s hard to say how likely this would be. Regardless, the entire scene is made up.
Honking loud
As with all the dinosaurs in PP, D. mirificus is shown making lots of sounds as it moves about its environment. We don’t know what sounds any dinosaur made, and while it is likely that therizinosaurs made some sounds, there is no reason to think that they were constantly broadcasting their whereabouts to the world.
That’s one big pile of…
All that food had to go somewhere, right? D. mirificus ends this scene by taking a massive dump in the water. This scene is funny and was definitely meant to be entertaining. The part that earns this scene it’s sin count comes next.
“And a giant such as Deinocheirus can produce over 20 tonnes of dung every year“
There is no quantitative evidence to support such a random (and large) number. We know that dinosaurs pooped, but we have no idea how much any dinosaur pooped.
Weird artistic choice
The shaggy coat of D. mirificus
We are shown a very shaggy D. mirificus here. As an ornithomimid we are pretty certain that it would have had simplistic protofeathers (van der Reest et al. 2016), but the level of shagginess shown here is not borne out from other ornithomimid specimens and certainly not from any D. mirificus fossils.
Speculative but nice to see
The terrible hand harvesting water plants
We are shown D. mirificus using its long arms to tear through the water plants in the swamp, collecting them up for easy access by the jaws. Although we have no data to support these motions, it is very nice to see a theropod using its arms for something. As we’ll see in future episodes, this is rarely considered.
Scene 4: South Africa

Baseless speculation
Cosmopolitan Quetzalcoatlus
This scene takes place in South Africa, yet we are told that the star animal of this scene, Quetzalcoatlus northropi can be found here. To Naish’s credit, he does address this point in the Twitter megathread by mentioning how researchers have posited that large pterosaurs were “continent hoppers”. While a fine bit of speculation, it still suffers from a lack of primary data (i.e., no Q. northropi fossils have been found in Africa).
Big eggs
The Q. northropi in this scene is a gravid female that has come to lay her eggs. Attenborough informs us that these large pterosaurs laid eggs that weighed up to 1 kg a piece. Can you guess how many azhdarchid eggs we have found to date? If you answered “none” then you would be correct. So, this number is just made up.
Delayed laying of soft-shelled eggs
We are shown the female laying a clutch of twelve, parchment-shelled eggs and then leaving them uncovered. Parchment-shelled, or “leathery” eggs are laid soft and then swell up and harden some as they gain moisture from their environment. This can’t happen if the eggs are just left out in the exposed air. They need to be buried. We are shown the female laying this clutch over the course of about a week. This is a weird strategy to employ for eggs that need to be buried for them to effectively take up moisture from the soil. We have no evidence to support this time-separated laying of eggs. This interpretation appears to be informed mostly from birds and their single oviducts, which is a weird choice since we have fossil evidence for paired ovaries in pterosaurs (Wang et al. 2015). A singular ovary / oviduct, appears to be a weird quirk of Aves alone (O’Connor et al. 2014). Birds lay hard-shelled eggs that remain exposed (typically covered only by the parents). Hard shells don’t wick moisture to the air the same way as parchment shells, so birds can afford this delayed laying strategy.
Mostly speculation
Parental care
Our female Q. northropi is shown making a nest and guarding it from intruders. This type of parental care is seen in a wide variety of sauropsids, so one could argue that there is precedence for this. That said, work on pterosaur embryos reveals that those studied were likely precocial (maybe even “super precocial), which would indicate reduced or nonexistent parental care in pterosaurs (Naish et al. 2021). For what it’s worth, a recent paper by Yang et al. (2023) has proffered potential evidence for more extensive (borderline altricial) parental care in Pteranodon ,though I’m not sure if one can really single out positive allometric growth from an animal that grew much larger than any of the other pterosaurs in the study. That’s a different story, though. For azhdarchids, as we don’t even have eggs from them we have no idea whether any of these nest guarding behaviours occurred.
The sound of thunder
Our mother to be is routinely shown making loud, booming calls. We have no real data on what pterosaurs sounded like, but this frequency range does at least align with the frequency ranges that we would expect them to hear well in based on inner-ear morphology.
Reasonable inference…but still speculation
Terrestrial Quetzalcoatlus
Attenborough informs the audience that Q. northropi is at home in the land and in the air, referring to some relatively extensive work done on terrestrial locomotion in azhdarchids (Witton and Naish 2008). The statement is hyperbolic as we don’t know just how “comfortable” these animals would be walking around, but given that some of the largest azhdarchids may have been secondarily flightless, this seems fine.
Leathery eggs
We are shown the mother laying twelve, soft-shelled eggs on the ground. This is a pretty solid inference based on our current knowledge of pterosaur egg anatomy, even if the exact number remains unknown.
Oophagous competitor
We see the rival female eating the eggs from the mother. This behaviour is documented in several extant sauropsids so it’s a reasonable inference even if we lack any primary data for it. The rival female could just have easily pushed the eggs aside, crushing them in the process.
Scene 5: Madagascar

Baseless speculation
Fuzzy Masiakasaurus with dental work
The scene opens with a beach full of modern-day crabs. Cretaceous crabs are known and some sub-fossil species have been recorded for Madagascar (Charbonnier et al. 2012), but I don’t think the showrunners worked that hard to nail down the type of crabs that would have been present here. We are introduced to Masiakasaurus knopfleri as it snatches up crabs along the beach. The animal is given a light coating of filaments and is festooned in fleshy lips. As a ceratosaur, M. knopfleri is far removed from any solid origin of filaments in dinosaurs. All ceratosaur fossils that retain any integument show scales and not filaments. In PP, as with T. rex, the show opted to go with a “compromise” that consists of a light coat of filaments across the body akin to what we see in elephants. As I’ve discussed several times before, this does not align with what we know of vertebrate integument development and largely relies on special pleading to achieve this look. It’s a borderline anti-scientific approach to reconstructing the dinosaur as it begrudgingly uses the primary data (fossils) and then shoehorns a preferred look for the animal.
A similar event seems to be occurring with the teeth. Now, the whole lip argument in theropods is one that I have largely stayed out of. I think it’s vastly overblown in its scientific importance (nearly the entire argument exists online) with opinions on lips being far stronger than the meager data we have one way or the other. Having said that, I find M. knopfleri to be a strange candidate for adding fleshy lips. A hallmark of this species is the weird tooth arrangement at the rostral end of the jaws (Sampson et al. 2001; Carrano et al. 2002). The teeth flair outwards in manner that remains difficult to reconcile with how the animal would have used them (e.g., dentary teeth 1–3 point forward from the jaw). The small but sharp teeth are indicative of piscivory, which aligns well with the environment where M. knopfleri is found. It’s possible that fleshy lips covered at least part of these strange teeth, but it’s equally possible that these teeth were better suited to a more gharial-like placement away from lips. Naish discusses this a bit in the Twitter Megathread.
Conventional reconstructions show these teeth radiating outwards and always obvious but with the jaw properly aligned and soft tissue added they just would not have been obvious when the jaws were closed, sorry! — Darren Naish
This is news to me as I’ve not seen any published reconstruction to this effect. Regardless of whether said teeth would have been exposed, their placement in PP does not reflect this unique arrangement. There are multiple instances in this scene where one can see the tooth arrangement up close and they do not show this outward angling at the rostral end of the jaw. They just look like bog-standard theropod teeth. I don’t find this reconstruction to be particularly accurate at all for this strange theropod.
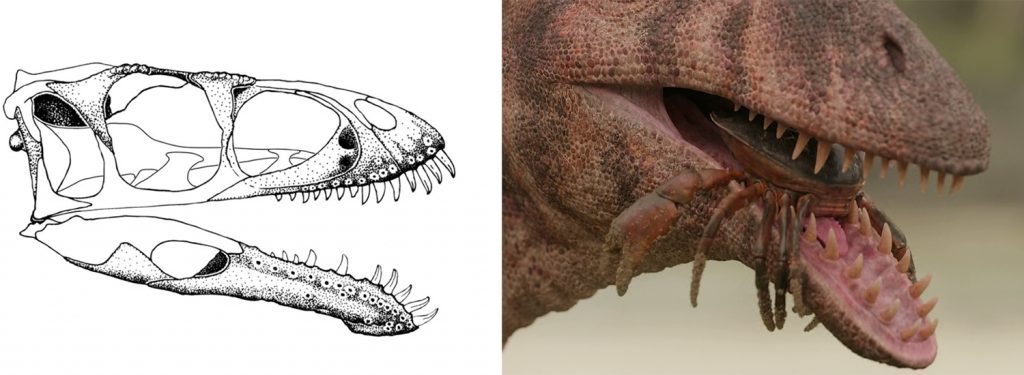
There are similar issues with the way we are shown M. knopfleri eating crabs. It just grabs them and gulps them down rather than moving them about in its jaws to crack into the shell. To be fair, there are no data on how M. knopfleri used its jaws, but if we turn to extant sauropsids that tackle crabs and other hard-shelled prey (e.g., Dracaena), there would be a greater degree of jaw manipulation and chewing than what we see here.
Doting parent
We are shown a set of three hatchlings covered in filaments (see, anti-science complaint above) rushing to mom for aid in eating food. The young are too small to tackle adult crabs by themselves and so rely on “their mother’s leftovers”. So, not only are we shown examples of extensive parental care in a species that we have only some ontogenetic data for, but it also includes parental feeding. Could some dinosaurs have fed their young? Sure. Do we have any way of telling if any dinosaur did this? No, not as of yet.
General complaints about hatchling dinosaurs
Aside from how the hatchlings look I could also live without the cartoony way in which they act. From constantly making noise to “acting” curious when looking at the crabs. None of this is how young sauropsids or even most young mammals act. It’s a very idealized version of what a “child” dinosaur would do rather than one based on the behaviours of extant hatchling and juvenile sauropsids.
Beezelbufo‘s eating habits
Our first introduction to Beezelbufo ampinga comes when it eats one of the hatchling M. knopfleri. After swallowing its prey, Attenborough informs us that B. ampinga “wont’ need to feed again for a month”. This is a completely made up number. It might be based on the way that extant pacman frogs (Ceratophrys), wherein they stay mostly sedentary during the dry season when prey is more sporadic. Whether B. ampinga experienced anything similar remains unknown.
Reasonable inference…but still speculation
Beezelbufo and Masiakasaurus
We are shown B. ampinga snatching up a young M. knopfleri. This would be a large prey item for a frog but definitely not beyond their capability. Extant bullfrogs (Lithobates catesbeianus) will ingest anything that they can fit in their mouths. B. ampinga is phylogenetically related to extant horned frogs (Ceratophryidae) which will also ingest anything they can fit in their mouths, so this is fully within the realm of possibility. The only things lacking are the primary data showing juvenile dinosaur parts within the stomach of B. ampinga.
New look, who dis?
The reconstruction of B. ampinga in PP is quite a bit different from previous restorations. As I mentioned above, B. ampinga is related to ceratophryids, which has led to most reconstructions showing it looking more like a typical horned frog. However, we don’t have any osteological correlates to verify this look. I was originally skeptical of this new look, but Naish did delve into this some more in part of the Twitter megathread. Though we may lack osteological correlates for the horned frog look, we can do a correlative comparison between skull shapes to at least argue a bit against the classic horned frog look.
Scene 6: Estuary

Reasonable inference…but still speculation
Brackish water visitors
There’s not much to this scene. It’s just a relaxing example of a potential estuary habitat in the Maastrichtian. The “star” of this scene is non-specific elasmosaurid just referred to as “elasmosaurs” by Attenborough. A pod of the animals are shown visiting the brackish estuary to explore the area. There’s not much to add here. Estuaries make for an interesting interaction between saltwater and freshwater species. It’s possible that this elasmosaurid species lived in pods as shown here, and it’s even likely that they visited estuaries at least occasionally, as we have several fossils that support young / small elasmosaurids in brackish and even freshwater (see Campbell et al. 2021 for a good review of this).
Missed opportunity
Plesiosaur hunting
We are shown the elasmosaurids hunting fish in the mixing waters of the river and the sea, but its all from afar and with no real insight into how these long-necked predators hunted. There have been several studies that have attempted to address this (Wilkinson and Ruxton 2012; Noè et al. 2017; Troelsen et al. 2019) and it would have been nice to see PP try to tackle some of this here rather than keep it in the dark and only show the occasional (and often overdone) breaching event.
In fairness to the T. rex roaring when making a kill, it might be trying to scare away scavengers that might try to steal from it. It would have been nice if the narrator explained it though.