
Most folks who visit my site by now have seen the big dinosaur news that has hit the interwebs. A new study from Matthew Baron, David Norman and Paul Barrett from University of Cambridge and the Natural History Museum of London, has seriously challenged the classic interpretation of dinosaur phylogeny.
Classical dinosaur phylogenetics
Although originally thought of as two unrelated branches of Reptilia that grew to immense size during the Mesozoic (e.g., Charig et al. 1965), for the last 43 years the group, Dinosauria, has been considered monophyletic (i.e., sharing a single origin) with the subgroups, Saurischia & Ornithischia, forming the first major branches within the group (Bakker et al. 1974). Saurischians, or “reptile hips” were aligned together by their similar hip shapes, skull characters (e.g., open antorbital fenestrae), and inferred soft tissues (e.g., air sacs). Ornithischians, or “bird hips” shared a hip structure that was superficially similar to that of birds, with a pubis that pointed caudally rather than rostrally, along with a variety of unique skull characters such as a neomorphic bone known as the predentary.
Study after study showed that this relationship was sound, and so it stayed that way. The problem with getting the same answer over and over again is that one eventually stops questioning it. Consistent results become common knowledge, and may even graduate to dogma. That’s not so bad if that common knowledge is true, but all too often many of these “obvious” cases wind up being just so stories upon closer inspection.
Throwing data at a classic relationship
Baron and company took a closer look at this classic relationship. They found that most studies supporting monophyly for Dinosauria, relied heavily on saurischians, with rarely more than one ornithischian as the representative for the “other team”. Similarly, these studies rarely included many examples of near dinosaurs, outgroup taxa that are believed to be close to the base of Dinosauria. Proper outgroups matter, and when you get closer to the base of an evolving lineage, they matter the most (since everyone basically looks the same at that point). Lastly, many of these studies used poor character choices for the ornithischian taxa. Taken together, these methods result in a phylogenetic analysis that is stacked heavily in favour of saurischians (theropods + sauropods) as a natural group.
In short, despite composing the majority of Dinosauria, ornithischians have consistently gotten the short end of the stick with dinosaur taxonomy.
Shredding the dinosaur family tree

Instead of looking at the popular, “showy” members of Saurischia and Ornithischia, Baron et al. focused on more basal members of each group. They looked at animals like Herrerasaurus ischigualastensis, Eoraptor lunensis, Fruitadens haagororum, Massospondylus (M. carinatus and M. kaalea),Agilisaurus louderbacki, Dracovenator regenti and 67 other taxa.
Along with this large list of taxa, Baron et al. used 457 different characters to score each taxon (character:taxon ratio of 6.2:1), with at least twenty of these characters being completely new. This was a large undertaking and took over three years to complete. So this should in no way be looked at as a quick “meta analysis” or generic “plug and chug” phylogeny. The authors went to great lengths to really assess the completeness and validity of their characters, as well as the robustness and stability of their trees.
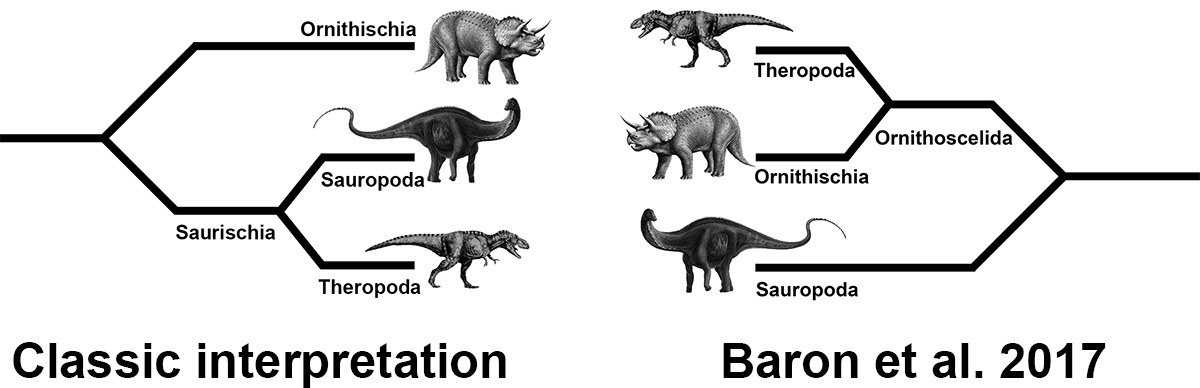
This is good, too, because their trees are bat shit insane compared to the classic interpretation of dinosaur phylogeny. The largest change that they found was that the “basally branching weird offshoots” that we call ornithischians, are actually more closely related to theropods than either group is to sauropods. Now it’s worth noting that Baron et al. were not the first to argue for closer ties between ornithischians and theropods. Thomas Henry Huxley first argued for this grouping back in 1870 (Huxley 1870). Indeed, Baron et al. resurrect Huxley’s original name for dinosaurs: Ornithoscelida (“Bird limbs”). Huxley considered sauropods to be closer to crocodiles than other dinosaurs, and so kept them out of his original Ornithoscelida definition.
Okay, so all that really happened is that Baron et al. moved Ornithischia deep into the Saurischia. On the outset that doesn’t seem like that big a deal. The biggest effect seems to be that ornithischians are no longer basally branching members of Dinosauria, and instead evolved from a saurischian ancestor. This actually jibes well with what we know of the ornithischian fossil record. Namely, that they are practically nonexistent until the early Jurassic. So what’s with all the hubbub?
A quick primer on phylogenetic taxonomy (PT) and phylogenetic nomenclature (PN)
During the late eighties, vertebrate paleontology started embracing a new way of describing evolutionary relationships. Instead of using the amount of shared characteristics as a means of determining who was related to whom, paleontologists were now using computers to run a basic model (maximum parsimony) across these laundry lists of characters. Maximum parsimony operates under the assumptions that new traits are rare and thus any evolutionary tree that produces the fewest number of new traits (steps), should be the correct one. This was the first time that phylogenetics could really be done with any degree of rigour. In the past, one basically relied on the professional opinions of people who spent their lives studying their animals of interest.
That was all fine and good, but along with this change to how phylogenetics was done, there was also a push for a new way of defining animal groups. In the past, one used the character diagnosis as the definition for the group. It made sense and it had the advantage of letting one quickly know what type of animal one was looking at by simply seeing if it checked off some / all of those laundry lists of characters. The disadvantage of this approach was that the list of diagnostic characters would change from analysis to analysis, requiring one to really pay attention to the latest work and to cite the appropriate author’s character definition. Apparently it was enough of a hassle that in 1986, vertebrae paleontologist Jacques Gauthier offered an alternative. Instead of defining a group by its diagnostic character set, define it in relation to the animals that its related to (Gauthier 1986). Gauthier along with Kevin de Quieroz (de Quieroz 1988, de Queiroz and Gauthier 1990) argued that this new approach to taxonomy was beneficial because it placed names firmly within the realm of evolution. In contrast, the Linnaean system of old was based on a time before widespread belief in evolution. This new approach was called: Phylogenetic Taxonomy (PT) or Phylogenetic Nomenclature (PN).
So for instance, Crocodylia (Crocodilia) used to be defined as reptiles that had the following anatomical characteristics:
- Broad, relatively long and flattened skull
- external nares (bony narial apertures) terminal
- fused frontal and parietal bones on skull
- antorbital fenestra reduced or absent
- highly developed secondary palate comprised of palatines and pterygoids
- Thecodont teeth
- Lack of prearticular and epipterygoid
- 9 cervicals
- Platycoelus caudal vertebrate (except for biconvex first caudal)
This definition comes from Romer’s Osteology of the Reptiles (1956) , and it is heavily truncated. The complete definition comprises a page and a half of text. Now, compare this to the modern, phylogenetic definition of Crocodylia as defined by Brochu (2003):
Last common ancestor of Gavialis gangeticus, Alligator mississippiensis,and Crocodylus niloticus and all of its
descendents [sic]
The latter definition is much quicker to digest and it anchors the taxonomic name of Crocodylia to animals that we have decided are true, “card carrying” crocodylians. The use of the last common ancestor approach brings evolution into it by ensuring that all animals descended from that ancestor are considered to be crocodylians. All of the characters listed in Romer’s definition are still used to form the group, but they no longer define it. The diagnosis has become separated from the definition.
However, you may notice that this definition is empty. It means nothing without foreknowledge of the relationship of the animals. One can’t just pick up a potential croc fossil in the field and determine that it’s a crocodylian without first inputting it into a phylogenetic analysis. The hollowness of this way of defining taxa was one of the original complaints about using PN over standard taxonomic practices. Proponents of PN argue that the emptiness of the names ensures taxonomic stability.
…phylogenetic definitions acknowledge that common ancestry is fundamental to the existence of the things being named. Accepting this proposition provides the basis for’ stable and unambiguous phylogenetic meanings of taxon names in the context of changing ideas about the details of phylogeny, both in terms of relationships and in terms of the precise level at which particular characters exist as synapomorphies. This stability and clarity derives from the fact that, for any two organisms, common ancestry at some level is certain, whereas the precise content of a taxon and its diagnostic characters are not. — de Quieroz & Gauthier 1990
To put it another way: The components (taxa) of a clade may change, but the definition stays the same.
Other complaints about PN comes from some of its stricter rules about taxon naming. For instance, any new taxon name is only considered valid if it incorporates all of the members found within that group. That is to say, the group has to be monophyletic. Groups that excluded some members (paraphyletic groups) or groups that are comprised of multiple unrelated members (polyphyletic groups) are considered invalid under phylogenetic taxonomy. Hence why birds—which descended from dinosaurs, which in turn descended of an earlier group of reptiles—are considered to be both dinosaurs and reptiles. Under PN, birds are also considered to be fish, invertebrates, and bacteria. A similar phylogenetic taxonomic relationship can be traced back for mammals too.
So those are the basic rules of phylogenetic taxonomy, and Baron et al.’s new phylogeny provides a perfect example of what happens if we actually follow those rules.
Beautiful disaster
In 1993, Kevin Padian and Cathleen May gave Dinosauria, Saurischia and Ornithischia, phyogenetically-based definitions:
We propose to define Ornithischia as all those dinosaurs closer to Triceratops than to birds, and Saurischia as all dinosaurs closer to birds than to Ornithischia. Dinosauria is defined as all descendants of the most recent common ancestor of birds and Triceratops.
Since this publication, the phylogenetic definition of Dinosauria has been given a slightly more specific modifier (Sereno 2005), being the most recent common ancestor of Passer domesticus and Triceratops horridus. Though, ultimately, it doesn’t really matter what bird species is used since they are all (theoretically) equally as related to dinosaurs under this definition. Ornithischia and Saurischia saw similar specific modification, as discussed by Baron et al. (2017):
[Saurischia] The most inclusive clade containing Passer domesticus and Saltasaurus loricatus but not Triceratops horridus (Sereno 2005)
[Ornithischia] The most inclusive clade containing Triceratops horridus but not Passer domesticus or Saltasaurus loricatus (Sereno 2005)
One look at our new phylogenetic tree for Dinosauria, and one can see where the problems start popping up.
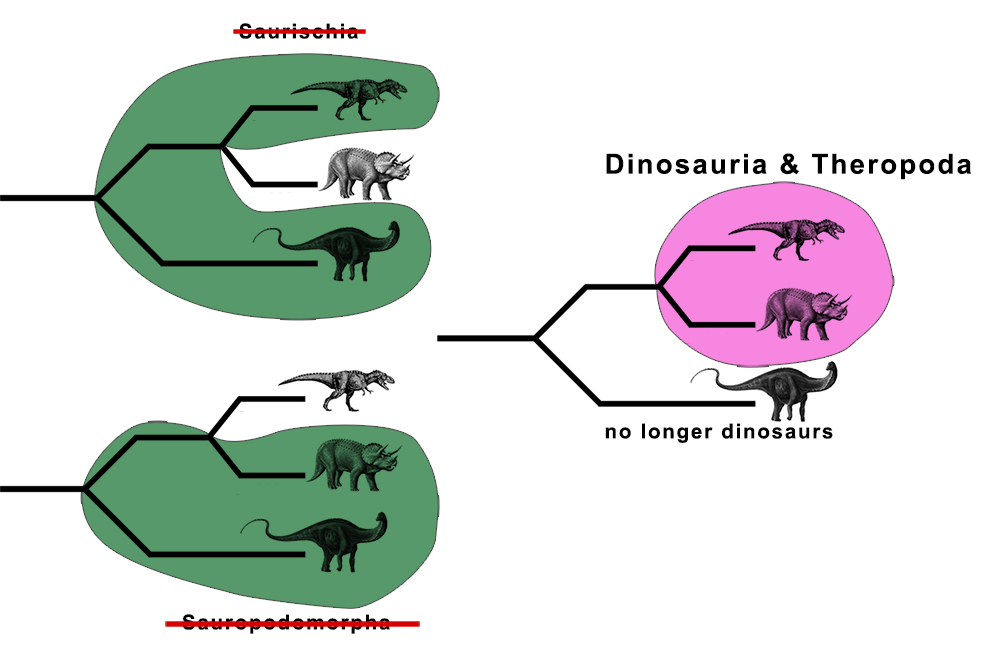
Moving ornithischians closer to theropods than either were to sauropods, essentially kicks sauropods out of Dinosauria completely. Similarly, Theropoda becomes synonymous with Dinosauria, given its phylogenetic definition. In fact, the clade that this new arrangement for Dinosauria encompasses, was once called Ornithoscelida, and is an older name than Theropoda. Thus, Theropoda should theoretically get sunk into Ornithoscelida.
Meanwhile, both Saurischia and Sauropodomorpha get booted out of existence due to their phylogenetic definition forcing a paraphyletic arrangement of taxa. So basically, all of the foundational taxonomic names for dinosaurs get completely reworked, and many animals that we have come to associate with theropods, sauropods, saurischians, and even Dinosauria itself, rearranged or (in the case of sauropods) thrown out. That’s a huge amount of taxonomic upheaval, and it’s contentious enough that Baron et al. couldn’t let it stand.
Redefining old names
Rather than simply explode dinosaur phylogeny and let the pieces land where they may, the authors opted instead to redefine many of these classic taxa so as to not lose the arrangements that are currently in place. The new definitions are as follows:
[Dinosauria] The least inclusive clade that includes Passer domesticus, Triceratops horridus and Diplodocus carnegii
[Theropoda] The most inclusive clade that contains Passer domesticus but not Diplodocus carnegii or Triceratops horridus
[Saurischia] The most inclusive clade that contains Diplodocus carnegii but not Triceratops horridus
[Sauropodomorpha] The most inclusive clade that contains Diplodocus carnegii but not Triceratops horridus, Passer domesticus or Herrerasaurus ischigualastensis
Now personally, I think this makes the most sense. Dinosauria and many of its subgroups, were named based on conceptual examples of the animals that contained them. Changing the affiliations up ultimately leads to unneeded confusion. So I can completely understand redefining these names so as to maintain their current and future usage.
However…that’s breaking the rules.
PN and the myth of stability
I’ve ranted about my feeling on phylogenetic nomenclature before. In short, I don’t like it. I think that the enforcement of monophyletic names onto well-established groups just causes unwanted confusion, heated arguments, and waters down any actual meaning. Not to mention the fact that it also necessitates the use of ugly modifiers just to talk about paraphyletic taxa (non-avian dinosaurs anyone?).

I also find the concept of divorcing diagnosis from definition is pointless, since diagnosis is often the best way to define something.
But still, what about those alleged advantages of PN? What about the stability of the names? Well, as Baron et al. 2017 have shown us, that is largely a lie. Despite the loud trumpeting from supporters of PN, that the constituents of a name are low priority compared to the stability of the system, this is yet another time where we see the rules of PN being broken in favour of keeping the constituent groups together. The redefinition of dinosaur groups by Baron et al. stands as prime example of why phylogenetics and classification are not the same thing and why they should not be treated the same way. We use phylogenetics to determine who is related to whom, but we use classification to determine what we call something. Both fields should work together to give us the most comprehensive picture of of life possible, but that doesn’t mean that one should be subservient to the other.
As I alluded to in the previous paragraph. This is not the first time that we have seen the rules of phylogenetic taxonomy bent to maintain standard taxonomic definitions for dinosaurs.
Back in 2011, Paleontologist Sterling Nesbitt published a landmark paper on the relationships of basal archosaurs (Nesbitt 2011). His analysis re-evaluated many poorly known taxa and tested the relationships of early archosaurs. His results found that the “near crocodiles” (Parasuchia), the phytosaurs, were not that closely related to any other archosaurs, and seem to be a sister group to all of Archosauria. The implications for this relationship were equally as disruptive as what we are now seeing with Baron et al. 2017.
Crurotarsi was previously defined as:
Ornithosuchidae, Parasuchia, Aetosauria, Rauisuchia, Crocodylomorpha, and all extinct descendants that
are most closely related to these taxa (Sereno and Arcucci, 1990)
Parasuchians (i.e., phytosaurs) are anchors to the clade name, Crurotarsi. Finding that parasuchians nested just outside of Archosauria means that all archosaurs are now nested within Crurotarsi, and as a result, dinosaurs are “crocodile-normal archosaurs” with a “crocodile-reversed” ankle. Now, Nesbitt did redefine Crurotarsi, Archosauria, and other archosaur groups, but these redefinitions were mostly a way to refine what the groups already meant. For instance, Crurotarsi was “redefined” as:
The least inclusive clade containing Rutiodon carolinensis Emmons, 1856, and Crocodylus niloticus Laurenti, 1768
So the arrangement of animals hasn’t changed, only the way in which the definition has been worded. Nesbitt stuck by the rules of PN, which means that dinosaurs are crurotarsans. The response from the paleocommunity has been interesting. There has been very little backlash to the findings, and most people seem to begrudgingly accept the new arrangement of taxa within Crurotarsi, but rather than going around talking about poposaurs and aetosaurs as non-ornithodiran crurotarsans, many in vert paleo systematics seem to have quietly just sunk Crurotarsi as a group name in favour of the older (but not synonymous) term: Pseudosuchia.
[Pseudosuchia] The most inclusive clade containing Crocodylus niloticus Laurenti, 1768, but not Passer domesticus Linnaeus, 1758 (sensu Sereno, 2005) — Nesbitt 2011
Now, six years after publication, there is nary a single paper referring to any Pseudosuchian as a non-ornithodiran crurotarsan. About the only thing brought up online about this change, was on the Chinleana website, along with some minor rumblings on the Dinosaur Mailing List. As with Baron et al. (2017), some of these conversations mentioned redefining Crurotarsi in a way that would keep crocs and dinosaurs split. It’s still possible that this might happen in the future. Though if it does, it will be breaking the rules of PN again.
So why is this a double standard?

Because, for many in dinosaur paleo, the argument for calling birds dinosaurs, or calling birds reptiles, is that it better reflect evolutionary relationships between these two groups, and it is the only way to maintain a phylogenetically consistent definition (i.e., it remains monophyletic). Because birds descended from a group of theropods, they have to be considered dinosaurs if Dinosauria is to be considered valid. This bias against paraphyly tends to be very strong.
Yet, if the reverse happens, and either suddenly dinosaurs get grouped into a different set of animals, or if not all classic dinosaurs are considered to be dinosaurs, there seems to be an agreed upon need to either redefine the definitions (thus eliminating the consistency of naming argument) or replacing the old name with a different, more exclusive name that does incorporate the animals of interest.
That said, it’s worth noting that I have seen a few folks online in favour of maintaining the old definitions and just dealing with the fact that sauropods would no longer be dinosaurs. However, the SAND (Sauropods Are Not Dinosaurs) proponents seem to be pretty few and far between. Nonetheless, kudos to standing by the rules of the preferred naming system.
I have also seen a few arguments put forth about the new definitions from Baron et al. not being valid under the PhyloCode, but that doesn’t really say much. PhyloCode has been in the works since 2000. It has received extensive pushback and as of this writing it still has hardly any support. For all intents and purposes, it was an idea that died on the vine.
The future of Dinosauria
So where does this leave Dinosauria? At the moment, the new tree from Baron et al. 2017 is the most up to date. It’s the culmination of 3 years of work looking at 74 specimens and around 35,000 data points. It’s unlikely we’ll see much of a topology change in the next year or so (ignoring cases where people re-analyze the same data). The results from Baron et al. are very intriguing because they show us (once again) that when we get towards the base of where a bunch of disparate animals branch off, everyone looks samey enough that relationships become very difficult to sort out.
Lastly, it’s worth noting that the consensus tree from Baron et al. only has a bootstrap support for Ornithoscelida, of 66%. That’s very low (in general, you want a bootstrap score of at least 80%), meaning that this clade only appeared in 66% of the random character samplings. However, it is also pertinent to point out that Ornithoscelida had a Bremer support value of 4, meaning that it takes the removal of at least 4 different characters before that clade collapses. This is a very strong support value and it held true regardless of whether all taxa were included, or if some were excluded (Baron et al. 2017 supplement). Bremer support values offer a better test of a tree’s robustness than bootstrap or jackknife scores, which is reflected in their higher computational costs (since it has to test these character removals across all the known trees). So there seems to be very strong support for Ornithoscelida as a clade. Now it’s possible that in the future, better characters and taxa will change this arrangement, but even with the low bootstrap score, these data seem to make it abundantly clear that when it comes to the most basic relationships among dinosaurs, things are very messy.
The next few years should prove to be very interesting for dinosaur systematics
~ Jura
References
Honestly, from a superficial visual standpoint the connections between sauropods & theropods (or the larger ornithischians & theropods), solidly-founded as they may be, have seemed rather tenuous at times. As such, I can halfway see where the SAND types are coming from. That said, full marks to Baron & company for a long-overdue reassessment of Dinosauria’s basal contingent.
How much time did it take for this new generation of scientists to declare a duck is a dinosaur but a diplodocus is not? Imagine what will happen in the next 10 years. A snake will be a mollusc, humans will be plants and cows will be minerals! Anyone who tries to define groups by their phylogeny without being sure of it should reconsider his career choices. This new method is worse. This whole new generation is worse. They should be ashamed of themselves.