
For a primer on this blog series and an FAQ see here.
| Other Episodes |
||||
| Coasts | Deserts | Freshwater | Ice Worlds | Forests |
| Islands | Badlands | Swamps | Oceans | North America |
This episode features contributions from the following paleontology consultants:

- Victoria Arbour
- Steve Brusatte
- Alexander Farnsworth
- Scott Hartman
- John Hutchinson
- Robert Spicer
- Paul Valdes
- Mark Witton
- Darren Naish
Scene 1: Forests of South America

Baseless speculation
Austroposeidon the elephant analogue
The star of this scene is the Brazilian titanosaur, Austroposeidon magnificus. At 25 meters long, this was a sizeable sauropod. Yet, in this scene we watch as several A. magnificus knock down trees to get to their juicy leaves. With such a long neck, you would think that that alone would provide these dinosaurs with the reach needed to get to their food. Making sauropods act like short-necked elephants is weird. Last I checked we don’t see a lot of giraffes knocking trees over to get to their leaves.
Hearty appetite
Attenborough informs us that this herd of 5–7 individuals require up to 10 tonnes of food a day. This sin is a two parter. First, we only have fossil evidence of a single A. magnificus. So, we don’t know if it was a herd animal or if it lived alone. Second, where did this 10 tonne number come from?
According to Naish, this wasn’t just pulled out of nowhere but was based on work by Martin Sander and colleagues (the Bonn sauropod working group). In particular, Jürgen Hummel’s work on digestibility of plant material. According to the Twitter megathread, these sauropods were reconstructed as living in a herd of 20 individuals (we never see this, but I guess the editing was meant to convey the number) eating about 500 kg of plant matter a day. Given this breakdown, these numbers add up. I think the show should have provided an aerial shot of the entire herd, or Attenborough should have explicitly mentioned the herd size.
All that said, the numbers are still too high. According to the Sauropod Working Group, estimated dietary intake (calculated via very general estimates of resting metabolism) tops out at 394 kg for a 70 tonne, fully obligate-endothermic sauropod (Hummel et al. 2008). For an animal like A. magnificus, the lower estimate for 30 tonnes is a more likely maximum (209 kg). So, the estimates are still too high here, regardless of thermophysiology.
Picky eater
Attenborough informs us that the knocking down of trees is to get to the tender leaves at the tips of the branches. Yet, if we are to believe that A. magnificus was chowing down on 500 kg of plant matter a day, then it couldn’t afford to be a picky eater here. More likely in this situation would be to show the animal just hoovering up all the leaves on the tree, and even including some of the branches. Naish argues that all herbivores show a preference for young leaves, but without a reference for said preference, I don’t see how this was ever tested.
Tree pushing specialist?
As we watch an A. magnificus take down a tree, Attenborough informs us that they are able to do this due to their “8 inch thick breastbone”. Where did this come from? As discussed below, A. magnificus is known from a handful of mostly fragmentary vertebrae. There is no preserved sternum. So, why make this whole bit up? The megathread discusses a bit on sauropod rearing only (no sternum anatomy mentioned), with Naish discussing how titanosaurs seem built well for tripodal stances. This is news to me, as the biomechanics do not bear this out (Mallison 2011).
Mostly speculation
Reconstruction of Austroposeidon
A. magnificus. It’s the largest sauropod known from Brazil during the Late Cretaceous. It’s also known from only two cervical vertebrae, seven fragmentary dorsal vertebrae, one good dorsal vertebra, and a partial sacral vertebra (Bandeira et al. 2016). So, most of what we are seeing in this scene is based on inferences from other, better-preserved titanosaurs.
Scene 2: American caving

Baseless speculation
Herd living Triceratops
The star of this scene is a herd of Triceratops horridus traveling through the forest. The cardinal sin in this scene is the presence of a herd of T. horridus. Whereas evidence of herd or herd-like living in ceratopsians is not rare, all of this primary evidence comes from centrosaurs such as Centrosaurus and Pachyrhinosaurus. Once we swap over to chasmosaurs, bonebeds become much less frequent; and when we look at just Triceratops, we find that these fossils are always found alone…save for one exception.
This one exception comes from the discovery of a Triceratops sp. bonebed in Montana dubbed the “Homer Site” (Matthews et al. 2009). This bonebed consisted of three juvenile to subadultTriceratops. As ceratopsian bonebeds go, this is pretty paltry (e.g., Centrosaurus apertus bonebeds contain hundreds to thousands of individual animals; Ryan and Russell 2001). That all three individuals here represent young animals could indicate that juvenile Triceratops may have occasionally formed small social groups. However, a review of the Homer Site bonebed by Keenan and Scannella (2014) indicated that these three individuals likely died from several separate flooding events and that this apparent monospecific accumulation of animals was more likely the result of a “bloat and float” scenario.
Based on our current data, Triceratops appears not to have been a particularly social ceratopsian, thus making this a strange narrative choice for PP to make.
Cave divers
In this scene, the herd of T. horridus are eating plants that release a deadly toxin for the animals. To offset this toxicity, the herd enters a cave in search of medicinal clay. This entire scene is based on observational studies of a single population of elephants from Kenya’s Mount Elgon region. This population mines Kitum Cave for its salt and the scraps travel down the food web to other animals in the region.
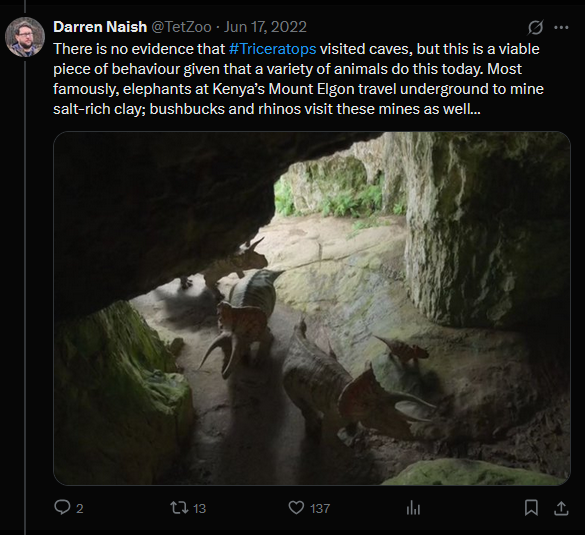
This is quite the unique observation, and its that uniqueness that is important. This is not something we observe in other elephants, nor in many animals outside of the region. In the megathread, Naish does offer several examples of animals that are entering caves for various reasons, but this one elephant population stands out for doing this salt mining. I don’t think it is a good idea to take a one-off example and just copy/paste it over to a very distantly related, extinct animal. I think the concept of showing such a massive animal traverse a cave is neat, but I don’t support the reasoning for it.
Good navigators
Our T. horridus herd are shown adeptly navigating the pitch black cave. This seems unlikely based on our current understanding of their paleoneurology (Sakagami and Kawabe 2020). A comparative paleoneurological look at the endocast of the brain in Triceratops found reduced olfaction and mediolateral eye stabilization. T. horridus may have just as likely barreled through structures rather than deftly move around them.
Mostly speculation…or maybe just an alternate perspective
Tighter packed forelimbs
The T. horridus herd are shown marching along with pretty tightly tucked forelimbs. According to the megathread, this was based on computer modeling of ceratopsian forelimbs. As far as I can tell (and based on consultants for this episode), these data likely come from Dempsey et al. (2023). These authors used the open source multibody dynamics software, GaitSym to test how estimated muscle placement and sizes would affect joint movements in several ornithischian taxa, including Triceratops. The results of their simulations found that ceratopsians, ankylosaurs, and stegosaurs all likely had a semi-sprawling posture to the forelimbs (flexed elbows, humerus jutting away from the body wall by ~32°). The authors rightly point out caveats to these hypotheses when using only muscle moment arms, but their results broadly agree with other work on ceratopsians (e.g., Thompson and Holmes 2007; Maidment and Barrett 2012). Yes, this paper did come out after the airing of Prehistoric Planet, but the analyses would have been going on at the time of production. So, I could see these data coming in this way.
So, why then is this herd of T. horridus shown with near parasagittal forelimbs and tightly tucked swings of their humeri? It may just be the angles shown, but I’m not seeing much forearm swing at all in these ceratopsians.
Scene 3: Forest in Patagonia

Baseless speculation
Male Carnotaurus on display
The star of this now infamous scene is Carnotaurus sastrei. The scene opens with a male taking a nod from bowerbirds and building a display space to show off to females. As of this writing we only have one skeleton of C. sastrei (Bonaparte et al. 1990). We don’t know anything about potential sexual dimorphism and we certainly don’t know anything about mating displays.
Ah, but things continue to get worse from here. Now that the male has captured the attention of the female he begins displaying to her…with his little arms.
Yes, this 2 tonne animal is dancing around this female showing off arms that are shorter than human arms. The arms are brightly coloured and he whips them around really fast, but they are still super tiny structures for an animal that would be better off using its large supraorbital horns. This is perhaps the most ridiculous bit of speculation that Prehistoric Planet entertains, and while it is baseless speculation I will give credit to the show for at least trying to justify it in this episode’s Prehistoric Planet Uncovered segment.
In the segment, Attenborough lays out how these arms were once thought of as vestigial remnants (spoiler alert: they still are) and that they were so small that the hands didn’t even have claws. We then cut to an interview with Prof. John Hutchinson who details the ball and socket style articulation of the humerus to the shoulder girdle and how it’s “weird”. I think this part was more done for entertainment purposes than anything else. Such a spherical humeral head is out of the ordinary for theropods, and aligns closer to humans, of all things. However, the size of the arms are what we should be keeping in mind here.
Attenborough later goes on to describe how the arm was heavily muscled. Indeed, one look at the shoulder girdle of C. sastrei and one could definitely get the impression that the arms were overmuscled. The coracoid in particular is very large. However, there are several neck and chest muscles that are attaching here too. The shoulder can still offer stabilizing functions that are unrelated to moving the arm (Burch 2017). That said, a comparative myological study of the arms in Majungasaurus crenatissimus (another, well-preserved abelisaurid) indicate that the arm muscles were capable of whipping the arm around rather quickly (Burch 2017) Distal attachment sites show reduced torque (weak arms) but the ability to produce large excursions for a given contraction. Burch posited that this motion could be useful for things like courtship and possibly display. Apparently, the PP team really jumped onto that passing statement about display.
Later in the Uncovered segment, Naish mentions that the “most likely explanation is that they were for display”. This is not the most likely explanation. The most likely explanation is that they were vestigial and that they may have been used for something. Display is a terrible use for such tiny structures. The video goes on to describe examples of animals using “highly specialized body parts that appear to have no other function” as mating displays. Their examples, though, highlight why C. sastrei and other abelisaurids were probably not using their arms for display. We are shown peacocks, birds of paradise, hummingbirds, and fan-throated lizards (Sitana). In every single case, the animals displaying are using large structures for display. If you are trying to impress the opposite sex or intimidate rivals, your display has to be large. There are no cases in which an animal uses a small structure for courtship display. That’s not how displaying works. So no, abelisaurids likely did not use their arms for display of any kind.

Instead, I would argue that courtship via tactile stimulation is the most likely hypothesis for these flexible yet tiny limbs. Because it turns out that a reduce limb with relatively large musculature and wide ranges of motion are exactly what we see in the spurs of pythons (Bellairs 1950).
Pythons and a few other basally-branching snakes retain aspects of their hindlimbs. This includes remnants of a pelvis, femur, and usually a nubbin representing the tibia (Bellairs 1950). The femur connects (or doesn’t) with the acetabulum of the pelvis (Palci et al. 2020) via a broad articulation that produces a very wide range of motion (at least a 112° fore-aft arc [Gillingham and Chambers 1982], with abduction from the body wall between 70°– 90° [Murphy et al. 1978]). Male pythons use their spurs during courtship to entice females, or help move them in position for copulation. No one is really sure exactly what function they perform during copulation, only that males use them during this time. Males scratch or rub their spurs on the female at a frequency of 1.6–3.5 Hz (Gillingham and Chambers 1982) and they do so with enough force to be heard up to 1.8 meters away (Murphy et al. 1978). If abelisaurids were using their teeny arms for anything, this would most likely be it.
As an aside I’ll never understand the fascination with vestigial forelimbs in theropods. We have allosaurians, ceratosaurians, megalosaurids, spinosaurids, and various coelurosaurs that all show well-developed arms, and yet one is hard pressed to find any studies on range of motion, grasping abilities, or other function aside from studies on flight. In contrast, T. rex and abelisaurs get the bulk of the arm studies. Seems like a lot of effort for arms that were obviously vestigial, even if they don’t fit the strict definition of “truly vestigial” (Smith and Wright 2022).
The rest of this scene is another copy/paste from what we see in Bower birds. Again, no evidence for this in abelisaurids, or any theropod.
Reasonable inference…but still speculation
The call heard around the forest
To get the attention of potential females, our protagonist emits and infrasonic boom from deep in his chest. Based on how the inner ear of dinosaurs are shaped (Gleich et al. 2005), and the general tendency for larger animals to hear at lower frequencies, an infrasonic boom is pretty reasonable. Crocodylians produce infrasound (Dinets 2013), as do many ratites (Mack and Jones 2003). Low frequency calls have also been recorded for turtles (Ferrara et al. 2013). So, having C. sastrei emit these calls is fairly reasonable, even if we don’t have any primary evidence for it (including inner ear endocasts).
Scene 4: Upland forest in China

Baseless speculation
Up in the mountains
This scene takes us to a mountainous forest in China. The problem here is that we don’t have any fossil sites from mountainous regions anywhere. It’s one of the limitations of fossilization. So, while dinosaurs and other animals likely did live up in these upland areas, we don’t have any direct evidence for them.
Fuzzy Qianzhousaurus
One of the two star dinosaurs in this scene is Qianzhousaurus sinensis, the long-snouted tyrannosaurine (alioramine) that was nicknamed “Pinocchio rex” by the media. As is standard fare with PP at this point, we see the presence of a feathered mohawk in this species, despite it being deeply nested within Tyrannosauridae. So, standard borderline anti-scientific sin here.
Temperature forest
Attenborough informs us that this is a temperate forest. Our two star taxa here are Q. sinensis and Corythoraptor jacobsi. Both species are known from the Nanxiong Formation. Sedimentological studies indicate a dry, arid environment with a mean annual temperature of 22–27°C (Zhao and Yan 2000). So, based on the actual formation that these animals are from, it’s clear that PP was not pulling from this location. Instead, we are looking at an area closer to the Yixian Formation (mean annual temp of ~10±4°C; Amiot et al. 2011), though even this formation reveals evidence for a warm-hot, humid and subtropical to moderately temperate environment based on plant remains (Cui et al. 2015). So, much of this scene is imagined, as it would have to be anyway given the upland location.
Mostly speculation
Group of azure Corythoraptor
We are shown a group of C. jacobsi donning some bright blue plumage. The colour choice is interesting as it doesn’t seem to make a lot of sense for a forest dwelling animal. That’s not really the sin here. I just think it’s a weird artistic choice. The only sin here is the representation of these oviraptorosaurs as group living animals. We only have a single specimen of C. jacobsi to go off of (Lü et al. 2017). This doesn’t mean that they didn’t live in groups, only that we don’t have the direct evidence for it. We do have an bonebed for the oviraptorosaur, Avimimus portentosus, so we know that at least one species in this group was at least partially social (Funston et al. 2016). Whether that justifies extending this behaviour to other species in the group is a matter of choice at the moment.
There is a section in the megathread in which Naish mentions how the cranial crests of C. jacobsi weren’t for obvious sexual display as “Both males and females apparently have similarly shaped crests and so on…” , which is a strange statement to make when we only have a single specimen of C. jacobsi (Lü et al. 2017). I think there is some confusion here over the comparison to cassowary casques in that paper (which are the same between the sexes, but not between the species). As discussed in a previous post, we don’t have the sample size needed to determine, nor separate, sexual dimorphism from species recognition. All that said, none of this is brought up in the show, and Twitter threads don’t count as official offshoots of Prehistoric Planet, so I’m not counting this as a sin.
Qianzhousaurus the tiger
Q. sinensis is seen slowly stalking the C. jacobsi herd before making an unsuccessful ambush attack on them. This same style is repeated a little later when our female Q. sinensis uses a thunderstorm to mask her footsteps. The entire scene is very much inspired by how tigers, leopards, and jaguars hunt. I’m okay with it as I think it’s probably pretty close to how these theropods would have hunted.
However…we still don’t have any primary evidence for this, so…
Scene 5: Burning forest in North America
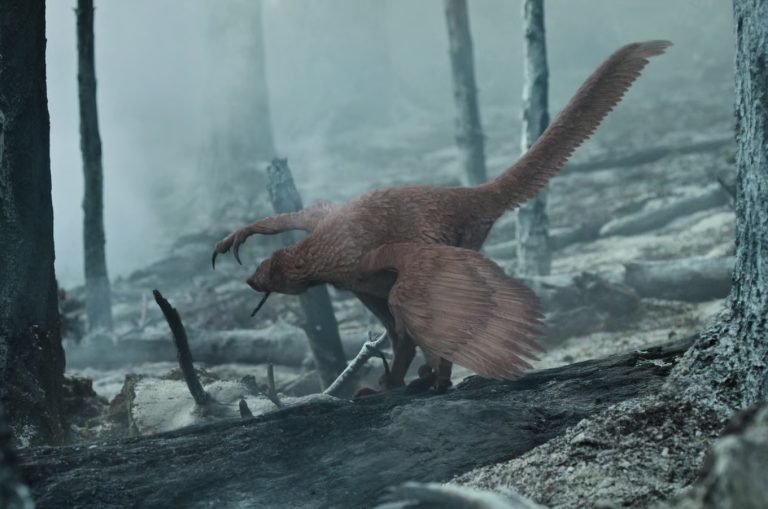
Baseless speculation
Golden eagle lookalike
The star of this scene is the dromaeosaurid, Atrociraptor marshalli. The animal is bedecked in a full plumage pattern reminiscent of extant Golden Eagles (Aquila chrysaetos). The choice of head to toe feathering is okay, as it lines up with how we understand the evolution of feathers in dinosaurs. Dromaeosaurids are a tough case as preserved data indicates that the tarsal and metatarsal region was scaled in some members of this group (Ji et al. 2001) and fully feathered in others (Xu et al. 2000). I can go either way on this. I would have liked to see a different colour scheme from just a A. chrysaetos copy/paste, but so be it. Aside from that, we have the same problem I mentioned in this old post regarding Jurassic World: Dominion. A. marshalliis not known from very much.
Smoking out parasites
The premise of this scene is that dinosaurs dealt with a lot of annual forest fires similarly to how extant animals do today. This is okay as we have sedimentological evidence for extensive wildfires in the Cretaceous (Brown et al. 2012, 2013) as well as some dietary evidence indicating that some herbivorous dinosaurs were eating charcoal from these fire deposits (Brown et al. 2020). The part where we fully enter into speculation is the use of smoke from forest fires to clean the wings of A. marshalli. As with the troodontid from Ice Worlds, this poor A. marshalli is risking burning its face by picking up smoking sticks to clean under its wings. To Naish’s credit, he states that this behaviour is completely speculative and based solely on a few species of corvids that are known to hold their wings over smoke.
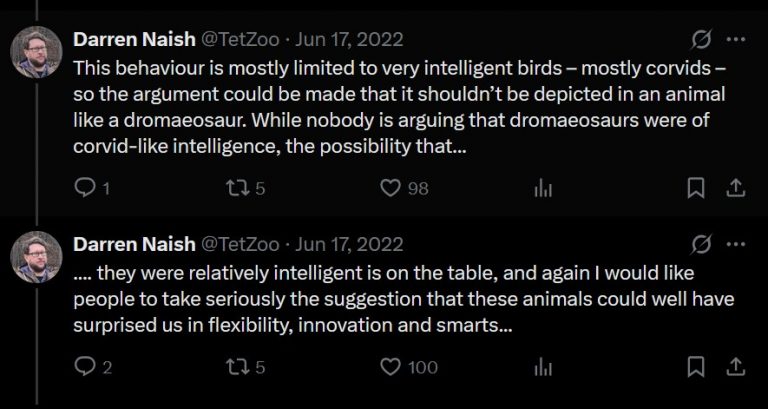
While I am fine with discussing how variable and sophisticated dinosaur behaviour may have been, I still find it irresponsible to apply unique examples like this without ever letting the audience know that we are just making things up.
Mostly speculation
Anodontosaurus chowing on charcoal
Later in this scene we see a large Anodontosaurus lambei moving through the recently burnt forest seeking out charcoal deposits to eat. This behaviour is based on a dietary study done for the nodosaurid, Borealopelta markmitchelli (Brown et al. 2020). As the authors noted in their discussion:
Any discussion of dietary reconstruction based on this specimen requires two caveats. Firstly, the data are based on a single specimen, which may not be representative of the species, or larger taxonomic groups as a whole. Secondly, the data represent a single brief event, probably of the order of hours, and at the end of one individual animal’s life, and may not accurately reflect the typical or average diet of the individual nor the taxon, especially in the context of seasonal changes and landscape variation in food availability.
So, transferring this well-documented last meal in a nodosaurid over to an ankylosaurid with a very different shaped snout probably doesn’t work. As other researchers have noted, these different snout shapes likely reflect different diets with ankylosaurids being “less picky” than nodosaurids (Mallon et al. 2014). That said, this still would not preclude A. lambei from searching out the occasional ash or charcoal deposit. We just don’t know.
Reasonable inference…but still speculation
A world burning
The B-Roll for the start of this scene is a series of forest fires. Attenborough informs us that these were annual occurrences. We don’t know if they were annual (we don’t have that kind of resolution for anywhere in the Mesozoic), but we have enough charcoal deposits to indicate that it was at least a fairly common event.
Scene 6: Night forest in Central Asia

Baseless speculation
Snoring sauropods
The scene opens at night where Attenborough tells us that “the night time forest is filled with strange sounds”. As the camera pans over, we see a large, sleeping sauropod. Attenborough informs the audience that the air sacs in the head and neck of sauropods, while weight saving, also amplify their snores.
That’s not how snoring works.
Mammals snore, and mammals may be the only animals that snore. Snoring is produced by the flapping of the soft palate against the roof of the mouth. Air that is partially blocked by a loose soft palate forces inhaled air to move in with greater force, causing tremors in the mucosa that ultimately produce the snoring sound (Vena et al. 2024). If you don’t have a soft palate that can get in the way of the glottis, then you can’t snore. Birds have a soft palate, but a glottis located at the base of the tongue, which slots right into the choana when the mouth is closed. Most squamates and testudines lack a secondary palate, meaning that air is free to reach the glottis when the mouth is closed (testudines reduce this further by turning their tongue into a secondary palate that provides direct access to their glottis). Those squamates that do have secondary plates (e.g., skinks) have bony secondary palates similar to crocodylians, which keeps anything from relaxing and getting in the way of air traversal. Dinosaurs fall somewhere in all of this mess. Exact palatal and choanal shape are hard to nail down but its enough to let us know that dinosaurs would not have snored.
And yes, that includes this cute little guy.
Despite the title and the narration, this little guy is not snoring. His syryingeal muscles (avian vocal folds) have relaxed enough to partially impede exhaled airflow and produce this melodic vibration. This is equivalent to a person moaning or lightly humming in their sleep.
No parents
The stars of this scene are a trio of young Therizinosaurus cheloniformis. We are shown these small juveniles sticking together as a group, sans any parents. This was an interesting take that I didn’t think too much about initially. There is an adult later in this scene but it’s not their parent. This is actually a nice change of pace from the overly doting parent trope we get in a lot of the PP scenes. We even have some partial support for this hypothesis based on the ossification pattern of embryonic therizinosaurids (Kundrát et al. 2008) However, it still should be said that we neither have hatchling nor juvenile T. cheloniformis material to go on, so it’s hard to say if this would be true for this species or not.
Climbing Therizinosaurus
Our trio of juveniles are roaming around at night in search of food. They find some potential food (a bee hive) up a tree and start climbing it. Attenborough informs us that “unlike most dinosaurs, young Therizinosaurus can climb”.
This is a strange statement to make. There isn’t anything special about T. cheloniformis anatomy to indicate arboreality nor unique scansoriality. They have long arms and scythe-like claws, but that doesn’t directly translate to climbing. According to the megathread, this climbing scene was meant to be a “call back” to earlier hypotheses of therizinosaur ecology that had them as tree climbing insect eaters.
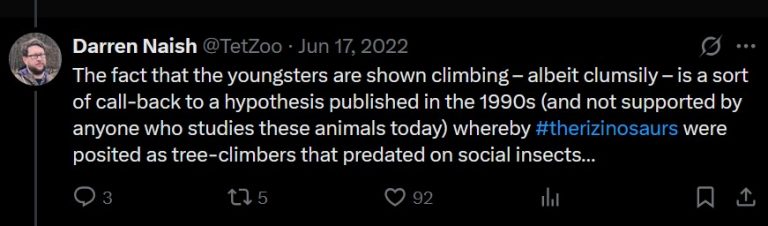
Did young T. cheloniformis climb trees at least on occasion? Sure, it’s plausible. Do we have any evidence to support this behaviour? No, not really.
Mostly speculation
Therizinosaurus growth rate
Attenborough informs us that these juvenile T. cheloniformis are only 1/10th the size of the adults at only six months of age. As far as I can tell we don’t have any growth rate data for therizinosaurs, so this is mostly made up. It’s hard to determine how much these juveniles have grown in six months when we aren’t sure what the starting size was.
Bee eaters
Our intrepid trio of juveniles discover a bee hive up a tree and proceed to eat from it. As Naish discusses in the megathread, this was another nod to older hypotheses of therizinosaur diets and ecology. Later, a large adult comes by and shreds open the hive to get at the honey inside.
Whether honey was a part of therizinosaur diets is largely dependent on whether or not honey existed in the Mesozoic. According to the megathread, we do have fossil evidence for honey-producing bees in the Cretaceous. I think that this may be going too far. We have one bee fossil from the Early Cretaceous Period, Melittosphex burmensis (Poinar and Danforth 2006). However, this bee looks a fair bit different from modern bees and shares several features that are transitional between bees and the wasp lineage they evolved from. Further, it is only a single specimen. No evidence is currently known for colonial living in this species. Aside from M. burmensis , the next oldest bee fossils come from the Paleogene period (Poinar and Danforth 2006), leaving a 35–45 million year gap in our knowledge of extinct bees. This one can go either way, though I would lean away from honey-producing social bees at the moment.
Scene 7: Island forest in Romania

Baseless speculation
Timid Telmatosaurus
The first animal we see in this scene is Telmatosaurus transsylvanicus, a basal hadrosauroid. Attenborough informs the audience that T. transsylvanicus seldom breaks cover. As this is a behaviour that doesn’t fossilize, we have no evidence to support this statement. Though small for its group, a T. transsylvanicus would still have been a fairly large animal. The narrative here seems to focus on the high foliage cover inferred for the Hateg Formation.
The size of Hatzegopteryx
We are next introduced to another enigmatic ornithopod, Zalmoxes robustus. We watch as a small group of these bizarre basal ornithopods run around the undergrowth avoiding the jaws of the azhdarchid, Hatzegopteryx thambena. Attenborough informs the audience that H. thambena stood 4.5 meters (15 ft) tall and was the heaviest animal ever to fly.
H. thambena is not known from very much. The holotype consists of some partial skull elements, and incomplete humerus and some unidentified bone fragments (Buffetaut et al. 2002). That’s not much to go on, but the humerus at least offers a hint at overall wingspan, suggesting an animal on par with Quetzalcoatlus northropi (Witton et al. 2010). There is also a referred femur (FGGUB R1625) with a length of 385 mm (lacking articular ends) which may belong to H. thambena and could be used to ballpark mass. According to Naish in the megathread, there is much more material for this species (or at least this genus) discovered, but none of it is published, which doesn’t help matters here.
To summarize, we have a good idea that H. thambena was a big azhdarchid, but we don’t have a lot of primary material to go on when making exact statements on height or mass.
Nuzzling titanosaurs
We eventually make our way towards a beach where we see a pair of Paludititan nalatzensis renewing their vows or something similar. The neck rubbing is very reminiscent of scenes out of The Land Before Time. I wouldn’t be surprised if this was inspired by that movie. All that said, we have no evidence for pair bonding in sauropods, or if neck rubbing was even a behaviour they performed. It’s certainly plausible that sauropods had submandibular glands akin to those seen in crocodylians and turtles (Mason and Parker 2010), but we don’t have any direct evidence for them at the moment. This type of neck nuzzling has a very avian quality to it, and was also likely inspired by that side of the dinosaur phylogenetic tree. Regardless, it remains purely speculative.
Mostly speculation
Salt lick
As all the dinosaurs introduced in the beginning of this scene make their way to the beach, Attenborough informs the audience that coating from the sea spray is a welcome source of salt for the herbivorous dinosaurs. We do know that several extant herbivorous animals will seek out salty structures that they can lick and/or eat when their diets are low in this electrolyte, but we don’t have any direct evidence that these island dinosaurs (most of Europe was island life back then) were doing this. I’m also unsure how prevalent such a salt search would be for animals living so close to saltwater.