
For a primer on this blog series and an FAQ, see here.
| Other Episodes |
||||
| Coasts | Deserts | Freshwater | Ice Worlds | Forests |
| Islands | Badlands | Swamps | Oceans | North America |

This episode features advisement by the following paleontology consultants:
- Victoria Arbour
- Steve Brusatte
- John Conway
- Alexander Farnsworth
- Scott Hartman
- John Hutchinson
- Robert Spicer
- Paul Valdes
- Mark Witton
- Darren Naish
Scene 1: Desert of South America

Baseless Speculation
Herd of Dreadnoughtus schrani males searching for females
We have no evidence for herding in D. schrani nor of any kind of sex segregation. We barely have evidence for the sex of any dinosaur. D. schrani is known from only two specimens collected from the Cerro Fortaleza Formation. This is a fluvial environment and not really a desert. D. schrani taphonomy suggests the animals were buried in an unexpected flooding event akin to river overflow as opposed to the flash flooding events characteristic of deserts. According to Naish, there were deserts “within walking distance” from where D. schrani is found. I’m not really sure what that is supposed to mean. Similarly, stating that lek-like gatherings must have happened is a bold statement. Behaviours seen in the present may have occurred in the past, but that is no guarantee that they did.
Hand claws…and hands
The two males that get into a fight are shown using their enlarged thumb claws to damage one another. However, a hallmark of titanosaurs is not only their lack of hand claws, but their lack of forelimb digits altogether (see Apesteguía 2005 for review). Only Diamantinasaurus is known to have any remnants of forelimb digits, with more advanced titanosaurs showing evidence for hyper-reduced and/or absent phalanges. It appears that this was based on an early misinterpretation of where Diamantinasaurus sat in the phylogeny of titanosaurs. If this part wasn’t highlighted in the show then I probably would have let this slide.
Inflatable neck displays
Easily the most far-fetched aspect of this episode. The male D. schrani are reconstructed with inflatable structures running alongside their necks. We know they are inflatable because we can see the males inflate them. Also, Attenborough states:
“Their hollow neck bones are connected to a series of bellows that inflate bizarre gular air sacs.”
There is zero evidence for this in the fossil record and even extant bases for this are extremely weak. It mostly boils down to “sauropods had air sacs, therefore…”. That’s not how air sacs work. This would require specialized tracheal air sacs that would go to inflatable throat sacs as in some chameleons (Huskey et al. 2018). This type of filling and emptying does not happen quickly and the exact mechanics behind how the sacs can inflate and deflate remain unknown. In contrast, birds like prairie chickens (Tympanuchus pallidicinctus) and frigate birds (Fregata) use esophageal air sacs (Riede et al. 2016), making their amazing mating displays equivalent to holding in a burp. To make something like this happen in an animal pushing 40 tonnes in mass would require some truly unique anatomy coupled with a standalone physiology (the air pressures needed to pull this off would not mix well with the rest of the soft tissues of the neck).
Rearing behemoths
I’m sticking this one in baseless speculation because of the choice of sauropod used for this. Biomechanical data lends some support to rearing in diplodocoids (compare Rothschild and Molnar 2005 with Mallison 2011). However for titanosaurs, not only is there no such support but there’s a fair bit of data to indicate that they would be the worst candidates for this (Mallison 2011).
The part that puts this in pure speculation land, though, is that they chose fully adult D. schrani for this behaviour. Animals that—according to the episode—are 40 tonnes in weight (the old, big male is given a near 50 tonne weight)! That is asking a whole lot for those hind limbs and tail to support, which is to say nothing of the force that the forelimbs would have to endure as the animal came crashing back down again. Big animals are less athletic than smaller animals. If these were young males at a third the size I would have been more forgiving, but not these two massive representatives of their species.
There’s also the issue of taking a head 12 meters above the ground and shooting it up another 5–7 meters without taking a massive hit to the animal’s blood pressure (more on this below). That the two males go at it again and again just makes the whole scene completely unrealistic.
Mostly Speculation
High browsing necks
I considered dropping this down to the “reasonable inference” level, but given that there was a strong narrative push to show all the sauropods as high browsers in this show, I feel this warrants the “mostly speculation” designation. Naish was part of a research group that challenged Kent Stevens’s work on sauropod neck posture (Taylor et al. 2009). This led to a short, vigorous, and mostly unpublished back and forth that was christened, the “neck wars” by the paleophile community. Personally, I think that the Stevens’s (2013) near point-by-point discussion of the counter-arguments proposed by Taylor et al. (2009) have yet to be fully resolved. There are some serious biomechanical and haemodynamic problems that have yet to be handled without keeping several sauropod necks rather horizontal. I also think it’s pertinent to point out that Stevens is not against necks held high for some sauropods. I don’t think that this is an all or none thing here and that there was probably enough diversity among sauropods to have a range of head heights.
But, that’s not what we get here. The narrative push was for high browsers only, as Twitter megathreads further bear out.
Reasonable inference…but still speculation
Sauropod walking
The limb movements shown for D. schrani as they walked around seem to lean on the data from Sellers et al. (2013) in which only one foot would move at a time. Now, during the making of this episode a more recent study came out to challenge some of these movements (Lallensack and Falkingham 2022), but since one can’t predict the future it’s understandable why it wasn’t used.
Hissing
Lots of hissing is heard from the sauropods as they both fight and display with one another. Lots of reptiles and birds hiss, so the precedent is there, it’s just hard to prove since the anatomical means of hissing don’t preserve in the fossil record. All that said, most sauropsid hissing is via a closed mouth rather than open. So…there’s that I suppose.
Scene 2: Desert of Asia

Baseless speculation
Velociraptor and Tarbosaurus
The main characters in this scene are Velociraptor and Tarbosaurus bataar. I’m not sticking the species epithet on Velociraptor here as the issue is that these taxa didn’t overlap in time. Velociraptor was a late Campanian animal, whereas the younger T. bataar falls within the Maastrichtian requirements of the show.
According to the Twitter megathread, this was based on a (currently) unnamed velociraptorine, but:
If this is meant to be a documentary then you absolutely can say that this is an indeterminate velociraptorine. I would even accept “an indeterminate dromaeosaur that may be a new species of Velociraptor”. You know, justify the speculation.
Instead, we see science communication take another backseat to entertainment.
Lightly fuzzy Tarbosaurus bataar
As with the first episode, this is almost antiscientific at this point.
Pack (pair) hunting “Velociraptor”
We are shown a pair of “Velociraptor” as they hunt down an unnamed agamid that is mostly comprised of B-roll from other BBC documentaries (it’s a real lizard for most of the scenes). Attenborough tells us that this little lizard needs to be careful:
“Because Velociraptors often hunt together”
We have circumstantial (and controversial) evidence that Deinonychus antirrhopus hunted in any sort of group (Ostrom 1969; Roach and Brinkman 2007). We have no evidence that any velociraptorine did the same. In fact, the best preserved specimen that we have for Velociraptor is from the Fighting Dinosaurs, in which a solitary animal is found dying in a failed hunt / defense. I would have put this in the mostly speculation section as it is certainly possible that these dromaeosaurs could have lived and even hunted in pairs, but the “documentary” decided to tell us that they “often do”.
Mostly speculation
A group of T. bataar
We are shown a group ofT. bataar snoozing in the afternoon sun. As with the velociraptorine bit above, this type of social interaction is well within the purview of extant reptiles and we have some slim support for group interaction in the older Albertosaurus sarcophagus (Currie and Eberth 2010). However, we don’t have any evidence for this in T. bataar.
Reasonable inference…but still speculation
Lazy Tarbosaurus
As mentioned above, we see a small group of T. bataar just lazying about in the sun. This is a behaviour that can’t really be captured in the fossil record, but it aligns well with what we know both of what apex predators do most of the time (sleep) and with how most animals handle the heat of mid-day in desert environments. The only potential change I would add here is having the animals try to find some shade.
“Velociraptor” flight feathers never retract
Our indeterminate velociraptorines are shown with primary (flight) feathers coming off their arms. This part is fine as it has a good basis in the phylogenetic placement of dromaeosaurs among maniraptorans, and with the primary evidence from the fossils of quill knobs in at least one individual (Turner et al. 2007). However, when extant birds are not using their primaries, they have them retracted and out of the way so they don’t get damaged. Velociraptorines probably didn’t fly much aside from some light gliding and parachuting, but I would imagine that they would still have the capacity to pull these delicate feathers out of the way when not in use, else they would get messed up pretty often, reducing their effectiveness at…most of what has been proposed for these structures. The presence of deep anchors within the ulna would further support some form of muscular control similar to extant birds.
Bonus sin: Cheaping out on the other wildlife

I mentioned this a bit earlier, but one of the “supporting cast” in this scene is an unnamed agamid. As far as I can gather, the filmmakers used an extant desert agama (Trapelus mutabilis) for most of this scene. So, much of the lizard shots are of a real lizard. The interactions with the dinosaurs change over to a CG lizard that doesn’t always match up colour wise. However, the part that warrants a sin here is not the CGI. It’s the fact that the creators opted to just toss in a “generic lizard” to fill their need rather than try to accurately reconstruct a lizard from that time period (e.g., Priscagama gobiensis). It really speaks to the priorities of the show that any “classic” reptile ends up getting short shrift.
Scene 3: Mononykus olecranus

Baseless speculation
The facial disc
We are told a lot of speculative stuff regarding M. olecranus in this scene, but none more speculative than the “facial disc” idea.
To start off, we have no direct evidence for feathers in M. olecranus, and must rely on its close association with Shuvuuia deserti. Of the two fossils known for S. deserti, one retains extremely poorly preserved structures that were hypothesized to have been feathers or protofeathers (Schweitzer et al. 1999), but remain controversial (Saitta et al. 2018). The point here being that our evidence for the type of filamentous coat we should expect in Alvarezsaurids is very hypothetical and poorly resolved. So, the filamentous disc idea—which is a unique structure seen today only in a handful of derived birds such as barn owls (Tyto alba)—is a phenomenal reach that is nowhere near concrete data.
This facial disc concept appears to be an idea that spawned largely from paleo-art. The seeds of this idea likely stem from Choiniere et al. 2021, which attempted to determine paleo-ecological separation via osteological correlates for hearing and vision. One of the conclusions by the authors was that alvarezsaurids show inner ear (cochlear duct) and scleral ring morphologies that are consistent with a nocturnal predator. It just so happens that the length of the cochlear duct in alvarezsaurids was most comparable to barn owls. Barn owls have a facial disc comprised of feathers, so….
So, a misunderstanding between inner ear anatomy and outer ear anatomy has produced the facial disc that you see in this scene. Meanwhile, the real take home from Choiniere et al. 2021 was that dinosaurs evolved enhanced hearing and nocturnal specializations more often than birds have.
Relatedly, given how heavily this interpretation relies on the work of Choiniere et al., you would think the showrunners would have shown this hunt taking place at night instead of during the day.
Super long tongue
Just after we see M. olecranus open up a termite nest (full of just modern-day termites), we are then shown the animal shoot out a tongue so long that it would make Gene Simmons blush. According to Attenborough, M. olecranus has:
A flexible tongue twice the length of her head.
Once again we see a direct comparison to mammals being brought in here. Alvarezsaurids have forelimbs roughly comparable to what we see in pangolins (though the latter are substantially longer), so therefore they must also have used a very long tongue like a pangolin.
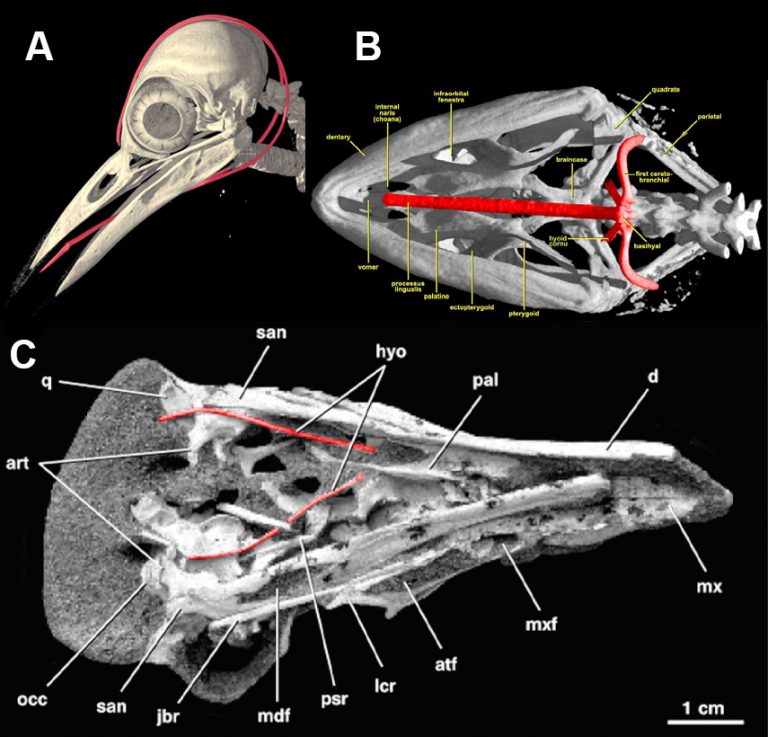
This association has no anatomical backing. Tongues leave marks on their respective bones, and the hyoid in particular is sculptured by its interactions to the extrinsic tongue muscles bound to it. Simplistic hyoids in most dinosaurs suggest unspecialized tongues (Li et al. 2018). To have a super long tongue, one needs to attach large/long protrusion muscles onto the hyoid somehow. This is perhaps most famously seen in chameleons and woodpeckers, which have notably enhanced hyoids. Woodpeckers take their ceratobranchials (hyoid horns) to a particularly crazy level and wrap them around their skull (Shufeldt 1900). Among the far more distantly related mammals, anteaters (Vermilingua) elongate their hyoids along with the rest of their skulls (Naples 1999), and pangolins (Pholidota) stand out from all other vertebrates in completely removing the tongue from the hyoid and migrating the tongue attachments onto the sternum (Chan 1995). It’s not easy to house a long tongue inside the body. Now, we do have a hyoid from M. olecranus (Chiappe et al. 1998) and it does appear more elongated than most other dinosaurs studied…but it doesn’t really stand out from a less-specialized maniraptoran such as Microraptor gui (see Fig. 8 of Li et al. 2018). We should expect to see similar anatomical changes in alvarezsaurids as we do in woodpeckers, chameleons, or any of the several other lizards that have long tongues, but we don’t. If they were nabbing stuff with their tongues then maybe they did it more similarly to horned lizards rather than woodpeckers.
Mononykus the bird
Throughout this scene we can hear M. olecranus regularly chirping away like a songbird. As with every dinosaur, we have no way of knowing what sounds any dinosaur made, but we do know that extant birds rely on their syrinx to generate sounds and we currently only find evidence for syringes up to the base of Neornithes (Clarke et al. 2016). So, it’s doubtful that M. olecranus was making these chirping noises unless it had convergently evolved a similar sound profile using laryngeal muscles (though those are also in doubt). That said, I’m mostly adding the sin based on this interview in which Naish mentioned working on making sure they didn’t just make the dinosaurs sound like extant animals. This one…just sounded birdy.
Mostly speculation
Social insect specialist
We are told that M. olecranus is a social insect specialist. Again, this interpretation is based largely on alleged fossorial adaptations in alvarezsaurids (namely, their ridiculously tiny arms). I’ve never understood why so many paleontologists are so fascinated by the vestigial arms of some dinosaurs, but never bother to study the well-developed arms of other dinosaurs. Regardless, we now have a handful of good biomechanical studies on alvarezsaurid forelimb range of motion, including two for M. olecranus itself. All bear out that forelimb use is most consistent with the motions of pangolin forelimbs. Pangolins are anteaters, so…
Yep, I did that again. As discussed by Choiniere et al. 2021:
Myrmecophagous mammals have previously been suggested as analogs for alvarezsauroids… However, we note that nocturnality, excellent hearing, and digging forelimbs occur much more widely in mammals that span a broad range of ecologies. Moreover, dental or mandibular reduction in mammals may not be a functional analog of dental or mandibular reduction in birds and other dinosaurs; extant birds with proportionally small crania or mandibles, such as galliforms, can have wide-ranging diets, and birds that consume small vertebrates can subdue their prey without the use of teeth. We therefore suggest that derived alvarezsauroids such as Shuvuuia could have consumed a wide range of nocturnal invertebrates and potentially small vertebrates, and that fossorially adapted forelimbs had versatile functions, allowing excavation of shallow-burrowing or crevice-inhabiting animals from the substrate or vegetation.
Whereas alvarezsaurids may have gone around excavating termite mounds, there is currently no reason to think that they weren’t also chasing down and eating up little lizards and mammals too.
Scene 4: Mongolian desert oasis

Baseless speculation
Camel of the Cretaceous
This scene features the hadrosaur, Barsboldia sicinskii. The animals are shown traveling in a herd across the desert, wherein we are told:
They are long distance specialists and can go without water for an astonishing length of time.
We have no way of knowing the water retention abilities or general water economy of any dinosaur. So, this part is purely made up.
Drinking behaviour
I hadn’t thought too much on it, but since it came up in the Megathread, it’s worth mentioning that we have no idea how any dinosaur drank water. This may sound strange at first glance, but water drinking behaviours are highly variable across vertebrates. Each group seems to have reinvented this wheel differently. Turtles siphon water through their mouths (and sometimes their noses), many birds do a scoop and toss method, cats use capillary action from their tongues, dogs toss water directly in their mouths, and snakes use a weird sponge action with their mouths. I’m sure that dinosaur drinking behaviour was equally variable. It’s just that we don’t know how to infer that behaviour as of yet.
Mostly speculation
The Mongolian titan
Our herd of B. sicinskii make it to an ephemeral pool where they are joined by some unnamed pterosaurs and a massive titanosaur known only as the “Mongolian titan”.
This unofficial name is based on a real fossil…well, a trace fossil. It’s a big footprint. Prehistoric Planet gave us a Cretaceous Bigfoot. 🙂
Of course, we are told that they are one of the largest animals on Earth, with a mass closing in at 70 tonnes. Needless to say, we can’t estimate any of that with any certainty from a single footprint.
Reasonable inference…but still speculation
Herds of B. sicinskii
This species is known from a single, fragmentary specimen, thus requiring any inferences of sociality to lean heavily on what we see in closer relatives. We do see several hadrosaur species forming death assemblages, so it’s not unreasonable. It’s just not known for this species.
Scene 5: Barbaridactylus mating ground

Baseless speculation
Mating ground plateau
We are shown Barbaridactylus grandis congregating on a plateau in the middle of a desert. This is an interesting idea, but not one borne from any known fossil locality.
Head crest
B. grandis is only known from a partial mandible and a handful of limb elements (Longrich et al. 2018). We have no cranial material to inform us on what the head of these animals looked like. The choice to run once more with the antler head crest was done to showcase sexual display structures in pterosaurs.
Mating scenes and sneaker males
This entire section feels heavily inspired by albatrosses, including the “beak jousting”. We have males battling for territory over females, sneaker males bypassing dominant males to mate, and females being generally promiscuous. All of these concepts have roots in extant animal observations, but again we have only a handful of pieces for B. grandis and no real data on mating systems in any pterosaur. Something like this may have happened in one or a few species, but we currently have no way of knowing if it did.
Aerial dogfight
Two males go at it in the air, resulting in a dynamic aerial battle worthy of Top Gun. In the end, one male sends another to his death as he tumbles from the sky. As with everything in this scene, we have no evidence to support any of these actions. I would further add that I’m not even sure that the male would have died from the height he fell from. Pterosaurs are famously light for their size, and even with a spiralling fall, I’m not sure their terminal velocity would have been high enough to be fatal. If B. grandis weighed 4 kg at adult size (pulled from Wikipedia, so be doubtful), then even a 1 km height would not be enough to break bones. There could be other crushing damage to soft tissues, including lethal concussions, though. It’s also possible that the acceleration observed is higher than gravitational acceleration. I don’t know, it leaves some interesting things to think about, but it’s still ultimately a made up story.
Scene 6: Gypsum desert of South America

Baseless speculation
Hadrosaur gastric constitution
We are brought to a gypsum desert in Argentina. This is likely based on a known evaporite deposit from that region. The star dinosaur this time is Secernosaursu keorneri. There is a bit of confusion here, as Naish mentions that their chosen taxon was changed to Huallasaurus after they had finished production. As far as I can tell, this name change was for Kritosaurus australis (Rozadilla et al. 2021). S. keorneri was never in question and Secernosaurus is a monospecific taxon. I think there may be some confusion there.
Regardless, the issue here is the following statement from Attenborough:
Although they can survive on a poor quality diet…
We have no dietary information for South American hadrosaurids, and certainly not for S. keorneri. Work on Edmontosaurus has shown a fairly wide dietary range, but we cannot, and should not just wantonly apply this to every hadrosaur.
Wise old ones
The herd is low on water and rely on the memory of their older members to guide them towards water. This is very elephant-like. Aside from elephants, whales, and humans, I’ve not heard of any other animals that rely on others this way. I don’t believe anything like this has been observed in sauropsids. Now, to be fair, I suspect that the main reason why it hasn’t been observed is because no one is looking, but still, if we are meant to rely on extant observations, then using examples from a deeply nested group of mammals is probably not the best well to pull from.
Dew drinkers
Later, we see the herd of S. keorneri reach the coast, where water vapour from the sea condenses on their scales and then lick the freshwater off their bodies. This is a behaviour seen in a few different desert animals, but it is one that requires two big things to work.
-
- One must be colder than the water vapour (condensation follows the opposite path of evaporation)
- The amount of water that forms will only benefit a small-bodied animal.
S. keorneri was small for a hadrosaur, but it was still around 200 kg. That’s too big to benefit from any condensation that would form on their scales. Further, without careful countercurrent heat exchange occurring along the skin, they probably would have been too warm for much moisture to condense at all.
Celestial navigation
The hadrosaur herd travels by night and uses the night sky as a map for navigating the mostly featureless terrain of the desert. This has some basis in that we know of many animals that utilize celestial maps for navigation. Studies on celestial navigators seem to focus either on insects, birds, or marine mammals (Foster et al. 2018). Whether terrestrial vertebrates take advantage of a stellar map in the night sky (aside from humans) remains unknown. I don’t see why it wouldn’t work, but we currently lack good evidence for it in extant animals. So, placing this on extinct animals that we know far less about is a bit of a stretch.
Mostly speculation
Happy wanderers
We are told that the S. keorneri herd must migrate from place to place for the herd to find enough food to survive. This makes sense when viewed from a (mammal-centric) perspective of limited food supplies in deserts. However, we have had several studies that have come out over the twenty-five years that indicate that endemism may have been the more common form of dinosaur distribution (e.g., Sampson et al. 2010; Benson et al. 2012) . So, perhaps S. keorneri did migrate, but it lived in a group a third this size, or they were solitary. All of these hypotheses are up for debate.
Reasonable inference…but still speculation
Night roamers
We are told that the S. keorneri herd prefers to move at night. Given that this is an animal shown living in a particularly hot desert, this type of movement makes sense. However—as usual—we don’t have any data to support this one way or the other.
Hi 🙂 I finally started watching prehistoric planet, and stumbled across your blog while searching for the science behind the show. While I have a background in biology, I know very little about dinosaurs, and thus I am unable to tell the difference between those physiology/behaviours that are reasonable speculation (perhaps based on a specific fossil), and those that are less reasonable (perhaps vaguely based on extant animals that occupied similar niches, etc.). I am disappointed to learn that rather a lot was made up, much more than what I expected… however, I now get to watch an episode, raise my eyebrows at certain things (“did Mononykus REALLY have a long anteater tongue…..? Wow! How do they know that?”) and then I get to speed on over to your blog afterwards to find out what’s what. So thank you for this series! I’m sure I will learn a lot 🙂
It’s always good to see these installments getting some use by others. Some of my issues definitely fall into nit picky “inside baseball” territory, but at least having them out there helps people look twice at these programs.